Привет! Скоро лето, а значит, все второкурсники медвузов будут сдавать биохимию. Сложный предмет, на самом деле. Чтобы немного помочь тем, кто повторяет материал к экзаменам, я решил сделать статью, в которой расскажу вам о «золотом кольце» биохимии — цикле Кребса. Его также называют цикл трикарбоновых кислот и цикл лимонной кислоты, это всё синонимы.
Сами реакции я распишу в . Сейчас же я расскажу о том, для чего нужен цикл Кребса, где он проходит и в чём его особенности. Надеюсь, получится понятно и доступно.
Для начала давайте разберём что такое обмен веществ. Это основа, без которой понимание Цикла Кребса невозможно.
Метаболизм
Одно из важнейших свойств живого (вспоминаем ) — это обмен веществ с окружающей средой. Действительно, только живое существо может что-то поглощать из окружающей среды, и что-то потом в неё выделять.
В биохимии обмен веществ принято называть «метаболизм». Обмен веществ, обмен энергией с окружающей средой — это метаболизм.
Когда мы, допустим, съели бутерброд с курицей, мы получили белки (курица) и углеводы (хлеб). В процессе пищеварения белки распадутся до аминокислот, а углеводы — до моносахаров. То, что я описал сейчас, называется катаболизм, то есть распад сложных веществ на более простые. Первая часть метаболизма — это катаболизм .
Ещё один пример. Ткани в нашем организме постоянно обновляются. Когда отмирает старая ткань, её обломки растаскивают макрофаги, и они заменяется новой тканью. Новая ткань создаётся в процессе синтеза белка из аминокислот. Синтез белка происходит в рибосомах. Создание нового белка (сложного вещества) из аминокислот (простого вещества) — это анаболизм .
Итак, анаболизм — это противоположность катаболизму. Катаболизм — это разрушение веществ, анаболизм — это создание веществ. Кстати, чтобы их не путать, запомните ассоциацию: «Анаболики. Кровью и потом». Это голливудский фильм (довольно скучный, на мой взгляд) о спортсменах, применяющих анаболики для роста мышц. Анаболики — рост, синтез. Катаболизм — обратный процесс.
Точка пересечения распада и синтеза.
Цикл Кребса как ступень катаболизма.
Как связаны метаболизм и цикл Кребса? Дело в том, что именно цикл Кребса является одной из важнейших точек, в которой сходятся пути анаболизма и катаболизма. Именно в этом и заключается его значение.
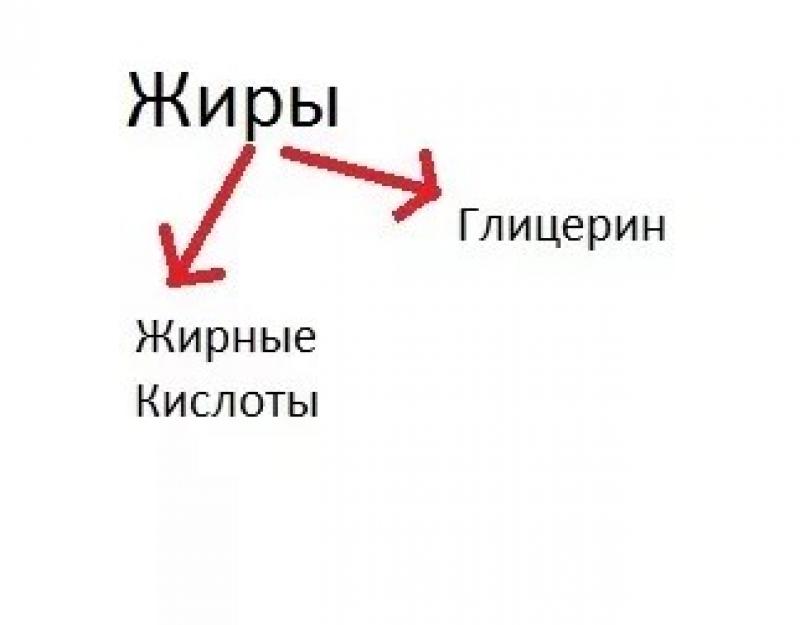
Давайте разберём это на схемках. Катаболизм можно условно представить как расщепление белков, жиров и углеводов в нашей пищеварительной системе. Итак, мы скушали пищу из белков, жиров, и углеводов, что дальше?

- Жиры — на глицерин и жирные кислоты (могут быть и другие компоненты, я решил взять самый простой пример);
- Белки — на аминокислоты;
- Полисахаридные молекулы углеводов — на одинокие моносахариды.
Далее, в цитоплазме клетки, последует превращение этих простых веществ в пировиноградную кислоту (она же — пируват). Из цитоплазмы пировиноградная кислота попадёт в митохондрию, где превратится в ацетил коэнзим А . Пожалуйста, запомните эти два вещества — пируват и ацетил КоА, они очень важны.
Давайте теперь посмотрим, как происходит этап, который мы сейчас расписали:

Важная деталь: аминокислоты могут превращаться в ацетил КоА сразу, минуя стадию пировиноградной кислоты. Жирные кислоты сразу превращаются в ацетил КоА. Учтём это и подредактируем нашу схемку, чтобы получилось правильно:

Превращения простых веществ в пируват происходят в цитоплазме клеток. После этого пируват поступает в митохондрии, где успешно превращается в ацетил КоА.
Для чего пируват превращается в ацетил КоА? Именно для того, чтобы запустить наш цикл Кребса. Таким образом, мы можем сделать ещё одну надпись в схеме, и получится правильная последовательность:

В результате реакций цикла Кребса образуются важные для жизнедеятельности вещества, главные из которых:
- НАДH (НикотинАмидАденинДиНуклеотид+ катион водорода) и ФАДH 2 (ФлавинАденинДиНуклеотид+молекула водорода). Я специально выделил заглавными буквами составные части терминов, чтобы легче было читать, в норме их пишут одним словом. НАДH и ФАДH 2 выделяются в ходе цикла Кребса, чтобы потом принять участие в переносе электронов в дыхательную цепь клетки. Иными словами, эти два вещества играют важнейшую роль в клеточном дыхании.
- АТФ , то есть аденозинтрифосфат. Это вещество имеет две связи, разрыв которых даёт большое количество энергии. Этой энергией снабжаются многие жизненно важные реакции;
Также выделяются вода и углекислый газ. Давайте отразим это на нашей схеме:

Кстати, весь цикл Кребса происходит в митохондриях. Именно там, где проходит и подготовительный этап, то есть превращение пирувата в ацетил КоА. Не зря кстати митохондрии называют «энергетическая станция клетки».

Цикл Кребса как начало синтеза
Цикл Кребса удивителен тем, что он не только даёт нам ценные АТФ (энергию) и коферменты для клеточного дыхания. Если посмотрите на предыдущую схему, вы поймёте, что цикл Кребса — это продолжение процессов катаболизма. Но вместе с тем он является и первой ступенькой анаболизма. Как это возможно? Как один и тот же цикл может и разрушать, и создавать?
Оказывается, отдельные продукты реакций цикла Кребса могут частично отправляться на синтез новых сложных веществ в зависимости от потребностей организма. Например, на глюконеогенез — это синтез глюкозы из простых веществ, не являющихся углеводами.
- Реакции цикла Кребса каскадны. Они происходят одна за другой, и каждая предыдущая реакция запускает последующую;
- Продукты реакций цикла Кребса частично идут на запуск последующей реакции, а частично — на синтез новых сложных веществ.
Давайте попробуем отразить это на схеме, чтобы цикл Кребса был обозначен именно как точка пересечения распада и синтеза.

Голубыми стрелочками я отметил пути анаболизма, то есть создания новых веществ. Как видите, цикл Кребса действительно является точкой пересечения многих процессов и разрушения, и созидания.
Самое важное
- Цикл Кребса — перекрёстная точка метаболических путей. Им заканчивается катаболизм (распад), им начинается анаболизм (синтез);
- Продукты реакций Цикла Кребса частично идут для запуска следующей реакции цикла, а частично отправляются на создание новых сложных веществ;
- Цикл Кребса образует коферменты НАДH и ФАДН 2 , которые переносят электроны для клеточного дыхания, а также энергию в виде АТФ;
- Цикл Кребса происходит в митохондриях клеток.
ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ (ЦИКЛ КРЕБСА)
Цикл трикарбоновых кислот впервые был открыт английским биохимиком Г. Кребсом. Он первым постулировал значение данного цикла для полного сгорания пирувата, главным источником которого является гликолитическое превращение углеводов . В дальнейшем было показано, что цикл трикарбо-новых кислот является тем центром, в котором сходятся практически все метаболические пути. Таким образом, цикл Кребса – общий конечный путь окисления ацетильных групп (в виде ацетил-КоА), в которые превращается в процессе катаболизма большая часть органических молекул , играющих роль «клеточного топлива »: углеводов , жирных кислот и аминокислот .
Образовавшийся в результате окислительного декарбоксилирования пирувата в митохондриях ацетил-КоА вступает в цикл Кребса . Данный цикл происходит в матриксе митохондрий и состоит из восьми последовательных реакций (рис. 10.9). Начинается цикл с присоединения ацетил-КоА к оксалоацетату и образования лимонной кислоты (цитрата ). Затем лимонная кислота (шестиуглеродное соединение) путем ряда дегидрирований (отнятие водорода ) и двух декарбоксилирований (отщепление СО 2) теряет два углеродных атома и снова в цикле Кребса превращается в оксалоацетат (четырехуглеродное соединение), т.е. в результате полного оборота цикла одна молекула ацетил-КоА сгорает до СО 2 и Н 2 О, а молекула окса-лоацетата регенерируется. Рассмотрим все восемь последовательных реакций (этапов) цикла Кребса .
Рис. 10.9. Цикл трикарбоновых кислот (цикл Кребса ).
Первая реакция катализируется ферментом цит-рат-синтазой, при этом ацетильная группа ацетил-КоА конденсируется с оксалоацетатом, в результате чего образуется лимонная кислота :

По-видимому, в данной реакции в качестве промежуточного продукта образуется связанный с ферментом цитрил-КоА. Затем последний самопроизвольно и необратимо гидролизуется с образованием цитрата и HS-KoA.
В результате второй реакции образовавшаяся лимонная кислота подвергается дегидратированию с образованием цис-аконитовой кислоты , которая, присоединяя молекулу воды , переходит в изолимонную кислоту (изоцитрат). Катализирует эти обратимые реакции гидратации–дегидратации фермент аконитатгидратаза (аконитаза). В результате происходит взаимоперемещение Н и ОН в молекуле цитрата :

Третья реакция , по-видимому, лимитирует скорость цикла Кребса . Изолимонная кислота дегидрируется в присутствии НАД-зависимой изо-цитратдегидрогеназы.

В ходе изоцитратдегидрогеназной реакции изолимонная кислота одновременно декарбоксилируется. НАД-зависимая изоцитратдегидрогеназа является аллостерическим ферментом , которому в качестве специфического активатора необходим АДФ . Кроме того, фермент для проявления своей активности нуждается в ионах Mg 2+ или Мn 2+ .
Во время четвертой реакции происходит окислительное декарбокси-лирование α-кетоглутаровой кислоты с образованием высокоэнергетического соединения сукцинил-КоА. Механизм этой реакции сходен с таковым реакции окислительного декарбоксилирования пирувата до ацетил-КоА, α-кетоглутаратдегидрогеназный комплекс напоминает по своей структуре пируватдегидрогеназный комплекс. Как в одном, так и в другом случае в реакции принимают участие 5 коферментов : ТПФ, амид липоевой кислоты , HS-KoA, ФАД и НАД + .

Пятая реакция катализируется ферментом сукцинил-КоА-синтета-зой. В ходе этой реакции сукцинил-КоА при участии ГТФ и неорганического фосфата превращается в янтарную кислоту (сукцинат ). Одновременно происходит образование высокоэргической фосфатной связи ГТФ за счет высокоэргической тиоэфирной связи сукцинил-КоА:

В результате шестой реакции сукцинат дегидрируется в фумаровую кислоту . Окисление сукцината катализируется сукцинатдегидрогеназой , в молекуле которой с белком прочно (ковалентно) связан кофермент ФАД. В свою очередь сукцинатдегидрогеназа прочно связана с внутренней ми-тохондриальной мембраной :

Седьмая реакция осуществляется под влиянием фермента фума-ратгидратазы (фумаразы ). Образовавшаяся при этом фумаровая кислота гидратируется, продуктом реакции является яблочная кислота (малат). Следует отметить, что фумаратгидратаза обладает стереоспецифичностью (см. главу 4) – в ходе реакции образуется L-яблочная кислота :

Наконец, в ходе восьмой реакции цикла трикарбоновых кислот под влиянием митохондриальной НАД-зависимой малатдегидрогеназы происходит окисление L-малата в оксалоацетат:

Как видно, за один оборот цикла, состоящего из восьми ферментативных реакций , происходит полное окисление («сгорание») одной молекулы ацетил-КоА. Для непрерывной работы цикла необходимо постоянное поступление в систему ацетил-КоА, а коферменты (НАД + и ФАД), перешедшие в восстановленное состояние, должны снова и снова окисляться. Это окисление осуществляется в системе переносчиков электронов в дыхательной цепи (в цепи дыхательных ферментов ), локализованной в мембране митохондрий . Образовавшийся ФАДН 2 прочно связан с СДГ, поэтому он передает атомы водорода через KoQ. Освобождающаяся в результате окисления ацетил-КоА энергия в значительной мере сосредоточивается в макроэргических фосфатных связях АТФ . Из 4 пар атомов водорода 3 пары переносят НАДН на систему транспорта электронов ; при этом в расчете на каждую пару в системе биологического окисления образуется 3 молекулы АТФ (в процессе сопряженного ), а всего, следовательно, 9 молекул АТФ (см. главу 9). Одна пара атомов от сукцинатдегидрогеназы-ФАДН 2 попадает в систему транспорта электронов через KoQ, в результате образуется только 2 молекулы АТФ . В ходе цикла Кребса синтезируется также одна молекула ГТФ (субстратное фосфорилирование ), что равносильно одной молекуле АТФ . Итак, при окислении одной молекулы ацетил-КоА в цикле Кребса и системе окислительного фосфорилирования может образоваться 12 молекул АТФ .
Если подсчитать полный энергетический эффект гликолитического расщепления глюкозы и последующего окисления двух образовавшихся молекул пирувата до СО 2 и Н 2 О, то он окажется значительно большим.
Как отмечалось, одна молекула НАДН (3 молекулы АТФ ) образуется при окислительном декарбоксилировании пирувата в ацетил-КоА. При расщеплении одной молекулы глюкозы образуется 2 молекулы пирувата, а при окислении их до 2 молекул ацетил-КоА и последующих 2 оборотов цикла трикарбоновых кислот синтезируется 30 молекул АТФ (следовательно, окисление молекулы пирувата до СО 2 и Н 2 О дает 15 молекул АТФ ). К этому количеству надо добавить 2 молекулы АТФ , образующиеся при аэробном гликолизе , и 6 молекул АТФ , синтезирующихся за счет окисления 2 молекул внемитохондриального НАДН, которые образуются при окислении 2 молекул глицеральдегид-3-фосфата в дегидрогеназной реакции гликолиза . Следовательно, при расщеплении в тканях одной молекулы глюкозы по уравнению С 6 Н 12 О 6 + 6О 2 -> 6СО 2 + 6Н 2 О синтезируется 38 молекул АТФ . Несомненно, что в энергетическом отношении полное расщепление глюкозы является более эффективным процессом, чем анаэробный гликолиз .
Необходимо отметить, что образовавшиеся в процессе превращения глицеральдегид-3-фосфата 2 молекулы НАДН в дальнейшем при окислении могут давать не 6 молекул АТФ , а только 4. Дело в том, что сами молекулы внемитохондриального НАДН не способны проникать через мембрану внутрь митохондрий . Однако отдаваемые ими электроны могут включаться в митохондриальную цепь биологического окисления с помощью так называемого глицеролфосфатного челночного механизма (рис. 10.10). Ци-топлазматический НАДН сначала реагирует с цитоплазматическим ди-гидроксиацетонфосфатом, образуя глицерол-3-фосфат. Реакция катализи-

Рис. 10.10. Глицеролфосфатный челночный механизм. Объяснение в тексте.
руется НАД-зависимой цитоплазматической глицерол-3-фосфат-дегидроге-назой:
Дигидроксиацетонфосфат + НАДН + Н + <=> Глицерол-3-фосфат + НАД + .
Образовавшийся глицерол-3-фосфат легко проникает через митохонд-риальную мембрану . Внутри митохондрии другая (митохондриальная) глицерол-3-фосфат-дегидрогеназа (флавиновый фермент ) снова окисляет глицерол-3-фосфат до диоксиацетонфосфата:
Глицерол-3-фосфат + ФАД <=> Диоксиацетонфосфат + ФАДН 2 .
Восстановленный флавопротеин (фермент-ФАДН 2) вводит на уровне KoQ приобретенные им электроны в цепь биологического окисления и сопряженного с ним окислительного фосфорилирования , а диоксиаце-тонфосфат выходит из митохондрий в цитоплазму и может вновь взаимодействовать с цитоплазматическим НАДН + Н + . Таким образом, пара электронов (из одной молекулы цитоплазматического НАДН + Н +), вводимая в дыхательную цепь с помощью глицеролфосфатного челночного механизма, дает не 3, а 2 АТФ .

Рис. 10.11. Малат-аспартатная челночная система для переноса восстанавливающих эквивалентов от цитозольного НАДН в митохондриальный матрикс. Объяснение в тексте.
В дальнейшем было показано, что с помощью данного челночного механизма лишь в скелетных мышцах и мозге осуществляется перенос восстановленных эквивалентов от цитозольного НАДН + Н + в митохондрии .
В клетках печени , почек и сердца действует более сложная малат-ас-партатная челночная система. Действие такого челночного механизма становится возможным благодаря присутствию малатдегидрогеназы и ас-партатаминотрансферазы как в цитозоле, так и в митохондриях .
Установлено, что от цитозольного НАДН + Н + восстановленные эквиваленты сначала при участии фермента малатдегидрогеназы (рис. 10.11) переносятся на цитозольный оксалоацетат. В результате образуется малат, который с помощью системы, транспортирующей дикарбоновые кислоты , проходит через внутреннюю мембрану митохондрии в матрикс. Здесь малат окисляется в оксалоацетат, а матриксный НАД + восстанавливается в НАДН + Н + , который может теперь передавать свои электроны в цепь дыхательных ферментов , локализованную на внутренней мембране митохондрии . В свою очередь образовавшийся оксалоацетат в присутствии глутамата и фермента АсАТ вступает в реакцию трансаминирования . Образующиеся аспарат и α-кетоглутарат с помощью специальных транспортных систем способны проходить через мембрану митохондрий .
Транспортирование в цитозоле регенерирует оксалоацетат, что вызывает к действию следующий цикл. В целом процесс включает легкообратимые реакции , происходит без потребления энергии, «движущей силой» его является постоянное восстановление НАД + в цитозоле гли-церальдегид-3-фосфатом, образующимся при катаболизме глюкозы .
Итак, если функционирует малат-аспартатный механизм, то в результате полного окисления одной молекулы глюкозы может образоваться не 36, а 38 молекул АТФ (табл. 10.1).

В табл. 10.1 приведены реакции , в которых происходит образование высокоэргических фосфатных связей в ходе катаболизма глюкозы , с указанием эффективности процесса в аэробных и анаэробных условиях
Министерство образования Российской федерации
Самарский Государственный технический университет
Кафедра «Органической химии»
Реферат на тему:
«ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ (ЦИКЛ КРЕБСА)»
Выполнил студент: III – НТФ – 11
Ерошкина Н.В.
Проверил.
Цикл Кребса
Цикл трикарбоновых кислот (цикл Кребса , цитратный цикл ) - центральная часть общего пути катаболизма , циклический биохимический аэробный процесс, в ходе которого происходит превращение двух- и трёхуглеродных соединений, образующихся как промежуточные продукты в живых организмах при распаде углеводов, жиров и белков, до CO 2 . При этом освобождённый водород направляется в цепь тканевого дыхания, где в дальнейшем окисляется до воды, принимая непосредственное участие в синтезе универсального источника энергии - АТФ .
Цикл Кребса - это ключевой этап дыхания всех клеток , использующих кислород, центр пересечения множества метаболических путей в организме. Кроме значительной энергетической роли циклу отводится также и существенная пластическая функция, то есть это важный источник молекул-предшественников, из которых в ходе других биохимических превращений синтезируются такие важные для жизнедеятельности клетки соединения как аминокислоты, углеводы, жирные кислоты и др.
Цикл превращения лимонной кислоты в живых клетках был открыт и изучен немецким биохимиком Гансом Кребсом , за эту свою работу он (совместно с Ф. Липманом) был удостоен Нобелевской премии (1953).
Стадии цикла Кребса
| Субстраты | Продукты | Фермент | Тип реакции | Комментарий | |
|---|---|---|---|---|---|
| 1 | Оксалоацетат + Ацетил-CoA + H 2 O |
Цитрат + CoA-SH |
Цитратсинтаза | Альдольная конденсация | лимитирующая стадия, превращает C 4 оксалоацетат в С 6 |
| 2 | Цитрат | цис
-акониат + H 2 O |
аконитаза | Дегидратация | обратимая изомеризация |
| 3 | цис
-акониат + H 2 O |
изоцитрат | гидратация | ||
| 4 | Изоцитрат + |
изоцитратдегидрогеназа | Окисление | образуется NADH (эквивалент 2.5 ATP) | |
| 5 | Оксалосукцинат | α-кетоглутарат + CO 2 |
декарбоксилирование | обратимая стадия, образуется C 5 |
|
| 6 | α-кетоглутарат + NAD + + CoA-SH |
сукцинил-CoA + NADH + H + + CO 2 |
альфакетоглутаратдегидрогеназа | Окислительное декарбоксилирование | образуется NADH (эквивалентно 2.5 ATP), регенерация C 4 пути (освобождается CoA) |
| 7 | сукцинил-CoA + GDP + P i |
сукцинат + CoA-SH + GTP |
сукцинилкофермент А синтетаза | субстратное фосфорилирование | или ADP ->ATP , образуется 1 ATP |
| 8 | сукцинат + убихинон (Q) |
фумарат + убихинол (QH 2) |
сукцинатдегидрогеназа | Окисление | используется FAD как простетическая группа (FAD->FADH 2 на первой стадии реакции) в ферменте, образуется эквивалент 1.5 ATP |
| 9 | фумарат + H 2 O |
L -малат | фумараза | H 2 O-присоединение (гидратация ) |
|
| 10 | L
-малат + NAD + |
оксалоацетат + NADH + H + |
малатдегидрогеназа | окисление | образуется NADH (эквивалетно 2.5 ATP) |
Общее уравнение одного оборота цикла Кребса:
Ацетил-КоА → 2CO 2 + КоА + 8e −Примечания
Ссылки
Wikimedia Foundation . 2010 .
- Цикл Кальвина
- Цикл Хамфри
Смотреть что такое "Цикл Кребса" в других словарях:
ЦИКЛ КРЕБСА - (цикл лимонной и трикарбоновой кислот), система биохимических реакций, посредством которой большинство организмов ЭУКАРИОТОВ получают основную энергию в результате окисления пищи. Происходит в КЛЕТКАХ МИТОХОНДРИЙ. Включает несколько химических… … Научно-технический энциклопедический словарь
цикл Кребса - Цикл трикарбоновых кислот, цикл последовательных реакций в клетках аэробных организмов, в результате которых происходит синтез молекул АТФ Тематики биотехнологии EN Krebs cycle … Справочник технического переводчика
цикл кребса - – метаболитический путь, приводящий к полному разрушению ацетил КоА до конечных продуктов – CO2 и H2O … Краткий словарь биохимических терминов
цикл Кребса - trikarboksirūgščių ciklas statusas T sritis chemija apibrėžtis Baltymų, riebalų ir angliavandenių oksidacinio skaidymo organizme ciklas. atitikmenys: angl. citric acid cycle; Krebs cycle; tricarboxylic acid cycle rus. цикл Кребса; цикл лимонной… … Chemijos terminų aiškinamasis žodynas
цикл Кребса - tricarboxylic acid (Krebs, citric acid) cycle цикл трикарбоновых кислот, цикл Кребса. Важнейшая циклическая последовательность метаболических реакций у аэробных организмов (эу и прокариот), в результате которых происходит последовательное… … Молекулярная биология и генетика. Толковый словарь.
ЦИКЛ КРЕБСА - то же, что трикарбоновых кислот цикл … Естествознание. Энциклопедический словарь
Цикл Кребса (Krebs Cycle), Цикл Лимонной Кислоты (Citric Acid Cycle) - сложный цикл реакций, где в качестве катализаторов выступают ферменты; эти реакции проходят в клетках всех животных и заключаются в разложении ацетата в присутствии кислорода с выделением энергии в виде АТФ (по цепи передачи электронов) и… … Медицинские термины
ЦИКЛ КРЕБСА, ЦИКЛ ЛИМОННОЙ КИСЛОТЫ - (citric acid cycle) сложный цикл реакций, где в качестве катализаторов выступают ферменты; эти реакции проходят в клетках всех животных и заключаются в разложении ацетата в присутствии кислорода с выделением энергии в виде АТФ (по цепи передачи… … Толковый словарь по медицине
ЦИКЛ КРЕБСА (цикл трикарбоновых кислот - цикл лимонной кислоты) сложный циклический ферментативный процесс, при котором в организме происходит окисление пировиноградной кислоты с образованием углекислого газа, воды и энергии в виде АТФ; занимает центральное положение в общей системе… … Словарь ботанических терминов
Цикл трикарбоновых кислот - Цик … Википедия
Цикл трикарбоновых кислот впервые был открыт английским биохимиком Кребсом. Он первым постулировал значение данного цикла для полного сгорания пирувата, главным источником которого является гликолитическое превращение углеводов. В дальнейшем было показано, что цикл трикарбоновых кислот является "фокусом", в котором сходятся практически все метаболические пути.
Итак, образовавшийся в результате окислительного декарбоксилирования пирувата ацетил-КоА вступает в цикл Кребса. Данный цикл состоит из восьми последовательных реакций (рис. 91). Начинается цикл с конденсации ацетил-КоА с оксалоацетатом и образования лимонной кислоты. (Как будет видно ниже, в цикле окислению подвергается собственно не ацетил-КоА, а более сложное соединение - лимонная кислота (трикарбоновая кислота). )
Затем лимонная кислота (шестиуглеродное соединение) путем ряда дегидрирований (отнятие водорода) и дскарбоксилирований (отщепление СО 2) теряет два углеродных атома и снова в цикле Кребса появляется оксалоацетат (четырехуглеродное соединение), т. е. в результате полного оборота цикла молекула ацетил-КоА сгорает до СО 2 и Н 2 О, а молекула оксалоацетата регенерируется. Ниже приводятся все восемь последовательных реакций (этапов) цикла Кребса.
В первой реакции, катализируемой ферментом цитратсинтазой, ацетил-КоА конденсируется с оксалоацетатом. В результате образуется лимонная кислота:

По-видимому, в данной реакции в качестве промежуточного продукта образуется связанный с ферментом цитрил-КоА. Затем последний самопроизвольно и необратимо гидролизуется с образованием цитрата и HS-KoA.
Во второй реакции цикла образовавшаяся лимонная кислота подвергается дегидратированию с образованием цис-аконитовой кислоты, которая, присоединяя молекулу воды, переходит в изолимонную кислоту. Катализирует эти обратимые реакции гидратации-дегидратации фермент аконитат-гидратаза:

В третьей реакции, которая, по-видимому, лимитирует скорость цикла Кребса, изолимонная кислота дегидрируется в присутствии НАД-зависимой изоцитратдегидрогеназы:

(В тканях существует два типа изоцитратдегидрогеназ: НАД- и НАДФ-зависимые. Установлено, что роль основного катализатора окисления изолимонной кислоты в цикле Кребса выполняет НАД-зависимая изоцитратдегидрогеназа. )
В ходе изоцитратдегидрогеназной реакции изолимонная кислота декарбоксилируется. НАД-зависимая изоцитратдегидрогеназа является аллостерическим ферментом, которому в качестве специфического активатора необходим АДФ. Кроме того, фермент для проявления своей активности нуждается в ионах Mg 2+ или Мn 2+ .
В четвертой реакции происходит окислительное декарбоксилирование α-кетоглутаровой кислоты до сукцинил-КоА. Механизм этой реакции сходен с реакцией окислительного декарбоксилирования пирувата до ацетил-КоА. α-Кетоглутаратдегидрогеназный комплекс напоминает по своей структуре пируватдегидрогеназный комплекс. Как в одном, так и в другом случае в ходе реакции принимают участие пять коферментов: TДФ, амид липоевой кислоты, HS-KoA, ФАД и НАД. Суммарно данную реакцию можно написать так:

Пятая реакция катализируется ферментом сукцинил-КоА-синтетазой. В ходе этой реакции сукцинил-КоА при участии ГДФ и неорганического фосфата превращается в янтарную кислоту (сукцинат). Одновременно происходит образование высокоэргической фосфатной связи ГТФ1 за счет высокоэргической тиоэфирной связи сукцинил-КоА:

(Образовавшийся ГТФ отдает затем свою концевую фосфатную группу на АДФ, вследствие чего образуется АТФ. Образование высокоэргического нуклеозидтрифосфата в ходе сукцинил-КоА-синтетазной реакции - пример фосфорилирования на уровне субстрата. )
В шестой реакции сукцинат дегидрируется в фумаровую кислоту. Окисление сукцината катализируется сукцинатдегидрогеназой, в молекуле которой с белком ковалентно связан кофермент ФАД:

В седьмой реакции образовавшаяся фумаровая кислота гидратируется под влиянием фермента фумаратгидратазы. Продуктом данной реакции является яблочная кислота (малат). Следует отметить, что фумаратгидратаза обладает стереоспецифичностью, - в ходе данной реакции образуется L-яблочная кислота:

Наконец, в восьмой реакции цикла трикарбоновых кислот под влиянием митохондриальной НАД-зависимой малатдегидрогеназы происходит окисление L-малата в оксалоацетат:

Как видно, за один оборот цикла, состоящего из восьми ферментативных реакций, происходит полное окисление ("сгорание") одной молекулы ацетил-КоА. Для непрерывной работы цикла необходимо постоянное поступление в систему ацетил-КоА, а коферменты (НАД и ФАД), перешедшие в восстановленное состояние, должны снова и снова окисляться. Это окисление осуществляется в системе переносчиков электронов (или в цепи дыхательных ферментов), локализованной в митохондриях.
Освобождающаяся в результате окисления ацетил-КоА энергия в значительной мере сосредоточивается в макроэргических фосфатных связях АТФ. Из четырех пар атомов водорода три пары переносятся через НАД на систему транспорта электронов; при этом в расчете на каждую пару в системе биологического окисления образуются три молекулы АТФ (в процессе сопряженного окислительного фосфорилирования), а всего, следовательно, девять молекул АТФ. Одна пара атомов попадает в систему транспорта электронов через ФАД, - в результате образуются 2 молекулы АТФ. В ходе реакций цикла Кребса синтезируется также 1 молекула ГТФ, что равносильно 1 молекуле АТФ. Итак, при окислении ацетил-КоА в цикле Кребса образуется 12 молекул АТФ.

Как уже отмечалось, 1 молекула НАДН 2 (3 молекулы АТФ) образуется при окислительном декарбоксилирова-нии пирувата в ацетил-КоА. Так как при расщеплении одной молекулы глюкозы образуются две молекулы пирувата, то при окислении их до 2 молекул ацетил-КоА и последующих двух оборотов цикла трикарбоновых кислот синтезируется 30 молекул АТФ (следовательно, окисление одной молекулы пирувата до СО 2 и Н 2 O дает 15 молекул АТФ).
К этому надо добавить 2 молекулы АТФ, образующиеся при аэробном гликолизе, и 4 молекулы АТФ, синтезирующихся за счет окисления 2 молекул внемитохондриального НАДН 2 , которые образуются при окислении 2 молекул глицеральдегид-3-фосфата в дегидрогеназной реакции. Итого получим, что при расщеплении в тканях 1 молекулы глюкозы по уравнению: C 6 H 12 0 6 + 60 2 -> 6СO 2 + 6Н 2 O синтезируется 36 молекул АТФ, что способствует накоплению в макроэргических фосфатных связях аденозинтрифосфата 36 X 34,5 ~ 1240 кДж (или, по другим данным, 36 Х 38 ~ 1430 кДж) свободной энергии. Другими словами, из всей освобождающейся при аэробном окислении глюкозы свободной энергии (окодо 2840 кДж) до 50% ее аккумулируется в митохондриях в форме, которая может быть использована для выполнения различных физиологических функций. Несомненно, что в энергетическом отношении полное расщепление глюкозы является более эффективным процессом, чем гликолиз. Необходимо отметить, что образовавшиеся в процессе превращения глицеральдегид-3-фосфата 2 молекулы НАДН 2 в дальнейшем при окислении дают не 6 молекул АТФ, а только 4. Дело в том, что сами молекулы внемитохондриального НАДН 2 не способны проникать через мембрану внутрь митохондрий. Однако отдаваемые ими электроны могут включаться в митохондриальную цепь биологического окисления с помощью так называемого глицерофосфатного челночного механизма (рис. 92). Как видно на рисунке, цитоплазматический НАДН 2 сначала реагирует с цитоплазматическим дигидроксиацетонфосфатом, образуя глицерол-3-фосфат. Реакция катализируется НАД-зависимой цитоплазматической глицерол-3-фосфат-дегидрогеназой.
Цикл Кребса также называется циклом трикарбоновых кислот , так как они образуются в нем в качестве промежуточных продуктов. Представляет собой ферментативный кольцевой конвейер, «работающий» в матриксе митохондрий.
Результатом цикла Кребса является синтез небольшого количества АТФ и образование НАД · H 2 , который далее направляется на следующий этап – дыхательную цепь (окислительное фосфорилирование), расположенную на внутренней мембране митохондрий.
Образовавшаяся в результате пировиноградная кислота (пируват) поступает в митохондрии, где она в конечном итоге полностью окисляется, превращаясь в углекислый газ и воду. Сначала это происходит в цикле Кребса, затем при окислительном фосфорилировании.
До цикла Кребса пируват декарбоксилируется и дегидрируется. В результате декарбоксилирования отщепляется молекула CO 2 , дегидрирование - это отщепление атомов водорода. Они соединяются с НАД.
В результате из пировиноградной кислоты образуется уксусная, которая присоединяется к коферменту А. Получается ацетилкофермент А (ацетил-КоА) – CH 3 CO~S-КоА, содержащий высокоэнергетическую связь.
Превращение пирувата в ацетил-КоА обеспечивает большой ферментативный комплекс, состоящий из десятков полипептидов, связанным с переносчиками электронов.
Цикл Кребса начинается с гидролиза ацетил-КоА, при котором отщепляется ацетильная группа, содержащая два атома углерода. Далее ацетильная группа включается в цикл трикарбоновых кислот.
Ацетильная группа присоединяется к щавелевоуксусной кислоте, имеющей четыре атома углерода. В результате образуется лимонная кислота, включающая шесть атомов углерода. Энергию для этой реакции поставляет макроэргическая связь ацетил-КоА.
Далее следует цепь реакций, в которых связанная в цикле Кребса ацетильная группа дегидрируются с высвобождением четырех пар атомов водорода и декарбоксилируются с образованием двух молекул CO 2 . При этом для окисления используется кислород, отщепляемый от двух молекул воды, а не молекулярный . Процесс называется окислительн ым декарбоксилирование м . В конце цикла щавелевоуксусная кислота регенерируется.
Вернемся на этап лимонной кислоты. Ее окисление проходит за ряд ферментативных реакций, при которых образуются изолимонная, щавелевоянтарная и другие кислоты. В результате этих реакций, на разных стадиях цикла, восстанавливаются три молекулы НАД и одна ФАД, образуется ГТФ (гуанозинтрифосфат), содержащий макроэргическую фосфатную связь, энергия которой впоследствии используется для фосфорилирования АДФ. В результате образуется молекула АТФ.
Лимонная кислота теряет два атома углерода с образованием двух молекул CO 2 .
В результате ферментативных реакций лимонная кислота превращается в щавелевоуксусную, которая снова может соединиться с ацетил-КоА. Цикл повторяется.
В составе лимонной кислоты присоединившийся остаток ацетил-КоА сгорает с образованием углекислого газа, атомов водорода и электронов. Водород и электроны переносятся на НАД и ФАД, которые являются акцепторами для него.
Окисление одной молекулы ацетил-КоА дает одну молекулу АТФ, четыре атома водорода и две молекулы углекислого газа. То есть углекислый газ, выделяемый при аэробном дыхании, образуется на этапе цикла Кребса . При этом молекулярный кислород (O 2) здесь не используется, он необходим лишь на этапе окислительного фосфорилирования.
Атомы водорода присоединяются к НАД или ФАД, в таком виде далее попадают в дыхательную цепь.
Одна молекула глюкозы дает две молекулы пирувата и, следовательно, два ацетил-КоА. Таким образом на одну молекулу глюкозы приходится два оборота цикла трикарбоновых кислот. В общей сложности образуются две молекулы АТФ, четыре CO 2 , восемь атомов H.
Следует отметить, что не только глюкоза и образующийся из нее пируват поступают в цикл Кребса. В результате расщепления ферментом липазой жиров образуются жирные кислоты, окисление которых также приводит к образованию ацетил-КоА, восстановлению НАД, а также ФАД (флавинадениндинуклеотида).
Если клетка испытывает дефицит углеводов и жиров, то окислению могут подвергаться аминокислоты. При этом образуются ацетил-КоА и органические кислоты, которые далее участвуют в цикле Кребса.
Таким образом неважно, каким был первичный источник энергии. В любом случае образуется ацетил-КоА, представляющий собой универсальное для клетки соединение.